Willkommen zu
Moderne Landwirtschaft
!


von Waldo G. Nuez-Ortín, Sam Ceulemans, Dolores I. Batonon-Alavo, Yves Mercier,
Adisseo
Pflanzliche Aquakulturfutter und Aminosäureergänzung
Fischmehl ist die bevorzugte Proteinquelle in Wasserfutterformulierungen. aber es wird immer weniger verfügbar und teurer. Billigere pflanzliche Proteinquellen wurden in der Futtermittelindustrie für Aquakulturen populär. insbesondere Soja- und Maismahlzeiten. Obwohl der Rohproteingehalt pflanzlicher Zutaten mit dem von Fischmehl vergleichbar sein könnte, solche Inhaltsstoffe sind im Allgemeinen weniger verdaulich und weisen einen Mangel an einer oder mehreren essentiellen Aminosäuren (EAA) auf. Bestimmtes, Aquafeeds auf Sojabohnenmehlbasis weisen einen Mangel an Gesamtschwefelaminosäuren (TSAA) auf und Aquafeeds auf Maismehlbasis weisen einen Mangel an Lysin (Lys) auf.
Außerdem, Der Mangel an TSAA nimmt mit steigendem Gehalt an Pflanzeninhaltsstoffen in Aquafeed-Formulierungen (Goff und Gatlin III, 2004). Daher, kristalline Aminosäuren, hauptsächlich Lys und Methionin (Met), werden zunehmend verwendet, um EAA-Mangel in pflanzlichen Aquafeeds zu füllen, um die Proteinsynthese und optimale Leistung aufrechtzuerhalten.
Da Met eine Vorstufe von Cystein (Cys) ist, die Konzentrationen an Nahrungs-Cys werden den Bedarf an Nahrungs-Met beeinflussen. Daher, diese beiden EAA zeigen überlappende Funktionen, und viele Studien bestimmen die TSAA-Anforderung (d. h. Met + Cys) anstelle der Met-Anforderung allein [National Research Council (NRC), 2011].
Obwohl sowohl die Met-Anforderung allein als auch die TSAA-Anforderung bei der Futtermittelformulierung berücksichtigt werden sollten, da sie zwischen den Arten stark variieren (siehe Tabelle 1; NRC, 2011), EAA-Anforderungen wurden im Allgemeinen unter optimalen experimentellen Bedingungen (d. h. Temperatur, Sauerstoffversorgung, verfügbares Essen, und Bevölkerungsdichte), es liegen jedoch keine Informationen zu den Anforderungen der EAA unter natürlichen Produktionsbedingungen vor.
Methionin-Supplementierung
Methionin kann in drei verschiedenen auf dem Markt erhältlichen synthetischen Formen ergänzt werden:L-Met (99 Prozent Wirkstoff), die biologisch aktive Form von Met in pflanzlichen oder tierischen Proteinen; DL-Met (99 Prozent Wirkstoff), eine racemische Mischung aus 50 Prozent D-Met und 50 Prozent L-Met, durch chemische Synthese hergestellt und jahrzehntelang als Futtermittelzusatz verwendet, und Hydroxymethionin (OH-Met), eine Hydroxylsäure mit einer Hydroxylgruppe (OH) anstelle einer Amingruppe (NH2) am asymmetrischen Kohlenstoff (siehe Abbildung 1). OH-Met ist entweder als Flüssigprodukt mit 88 Prozent Wirkstoff und 12 Prozent Wasser oder als Calciumsalz erhältlich.
Unterschiede in der chemischen Struktur führen zu Unterschieden in der Absorption, Wandlung, und Nutzung dieser Moleküle. Ähnlich wie bei anderen EAA, DL-Met wird über aktiven Transport aufgenommen. Im Gegensatz, weil OH-Met eine organische Säure ist, es wird sowohl durch passive Diffusion als auch durch aktiven Transport absorbiert (Martín-Venegas et al., 2007), die Na+/H+-abhängig ist und vom Monocarboxylat-Transporter 1 betrieben wird.
Außerdem, während sowohl D- als auch L-Met über den Magen-Darm-Trakt resorbiert werden, Die OH-Met-Resorption ist am Ende des Duodenums abgeschlossen (Richards et al., 2005; Jendza et al., 2011). Die Umwandlung von L- und D-OH-Met in L-Met ist ein zweistufiger Prozess, jede Verbindung wird zuerst in Ketomethylthio-Buttersäure (KMB) umgewandelt und dann in L-Met transaminiert (Vázquez-Añon et al. 2017) (siehe Abbildung 2). Diese Umwandlung wird durch verschiedene Enzyme im Magen-Darm-Trakt angetrieben, Leber und Niere (Fang et al. 2010).
Die Wirkung einer Met-Supplementierung geht über die Proteinsynthese hinaus. Der unterschiedliche Stoffwechsel von OH-Met trägt zur Verbesserung der antioxidativen Kapazität und des Immunsystems von Tieren bei, sowie zur Linderung von Stressreaktionen (Métayer et al., 2008). Das Konzept einer besseren antioxidativen Wirkung von OH-Met im Vergleich zu DL-Met ist mit seiner größeren Tendenz verbunden, am Transsulfurierungszyklus beteiligt zu sein. und größere Mengen an Cys zu bilden, Taurin und Glutathion (Martín-Venegas et al., 2006) sowie zur Erhöhung des reduzierten Glutathion/oxidiertes Glutathion-Verhältnisses (Willemsen et al., 2011).
Bei Knochenfischen, die schützende Rolle von OH-Met wurde mit der Taurinproduktion und mit der Verbesserung der physischen Barriere im Darm und in den Kiemen sowie des antioxidativen Status und der Immunität in Verbindung gebracht (Wu et al., 2018). Cys und Glutathion wirken als kettenbrechende Elemente der oxidativen Prozesse (Métayer et al., 2008). Ihre verstärkte Produktion durch OH-Met verbessert die Fähigkeit zum Abfangen von Hydroxylradikalen und die enzymatische antioxidative Kapazität. wodurch die Fleisch- und Filetqualität durch die Verringerung der Protein- und Lipidoxidation verbessert wird (Xiao et al., 2012; Lebret et al., 2018).
Ähnliche Biowirksamkeit von Methioninquellen bei Wassertieren
Trotz der geringen Zahl von Studien zum Vergleich von Methioninquellen bei Wassertieren, Mehrere Studien zeigen, dass Fische DL-Met oder OH-Met genauso effektiv verwenden können wie L-Met, um ihren TSAA-Bedarf zu decken. Da die Schätzung der Bioeffizienz (d. h. der biologischen Effizienz in Bezug auf die Wachstumsleistung wie Körpergewichtszunahme und Futterverwertungsverhältnis) von DL-Met und OH-Met nicht immer die Unterschiede in der Futteraufnahme berücksichtigt, In der Literatur finden sich widersprüchliche Ergebnisse.
Dietary Met beeinflusst die Futteraufnahme und damit die Leistung (Vázquez-Añón et al., 2017). Angesichts des engen Zusammenhangs zwischen den zirkulierenden Met-Spiegeln und der freiwilligen Futteraufnahme und der Tatsache, dass OH-Met als OH-Met und nicht als Met an die Gewebe abgegeben wird, dieser metabolische Unterschied könnte zu unterschiedlichen Fütterungsmustern nach Belieben zwischen OH-Met- und DL-Met-supplementierten Tieren führen (Vázquez-Añón et al., 2017).
Deswegen, die von einem Tier aufgenommene Met-Menge (Met-Aufnahme =Nahrungsmet x Futteraufnahme) sollte beim Vergleich von Met-Quellen berücksichtigt werden, da dieses Kriterium bei anderen Aminosäuren wie Lysin befolgt wurde (Smiricky-Tjardes et al. 2004). Unter dieser Überlegung, sowohl OH-Met als auch DL-Met waren zu 100 Prozent äquivalent zu L-Met (Agostini et al., 2015; Vázquez-Añón et al., 2017).
Unter den 54 von Experten begutachteten Studien, die die Biowirksamkeit der Met-Supplementierung berichteten, die von 1978 bis 2019 veröffentlicht wurden, 24 verglichen die Supplementation verschiedener Met-Quellen in unterschiedlichen Dosen mit einer Met-defizienten Ernährung, aber nur 17 dieser Studien wurden unter Bedingungen durchgeführt, die einen genauen Vergleich zwischen Met-Quellen ermöglichten.
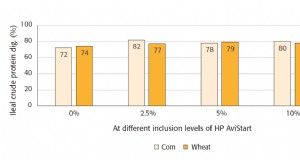
In diesen Studien, die verschiedenen Met-Quellen wurden auf gleicher Schwefel- oder äquimolarer Basis geliefert, um bei allen experimentellen Behandlungen ähnliche Mengen an Met + Cys zu erhalten, sowie ähnliche Mengen der anderen Nährstoffe. Die verschiedenen Met-Quellen führten zu ähnlichen Wachstumsleistungen. Zum Beispiel, Goff und Gatlin III (2004) berichteten von L-Met, DL-Met, und OH-Met als gleichermaßen wirksame Met-Quellen für Rote Trommel (Sciaenops ocellatus) (siehe Abbildung 3). Forster und Dominy (2006) stellten fest, dass diese Quellen auch verwendet werden könnten, um den Met-Bedarf von pazifischen weißen Garnelen (Litopenaeus vannamei) zu decken. Ma et al. (2013) fanden heraus, dass Steinbutt (Psetta maxima) OH-Met genauso effektiv einsetzen kann wie oder besser als, L-Met, um eine maximale Leistung zu erzielen.
Bei suboptimalen Nahrungsmengen, mit L-Met gefütterte Fische schnitten besser ab als mit OH-Met gefütterte Fische; im Gegensatz, Letztere schnitten in der Nähe des Erfüllten Anforderungsniveaus besser ab. Für Kanalwelse (Ictalurus punctatus), Zhao et al. (2017) fanden heraus, dass die Supplementierung von Met-Mangeldiäten mit OH-Met und mikroverkapseltem DL-Met, anstelle von kristallinem DL-Met, könnte die Wachstumsleistung dieser Art gleichermaßen verbessern (1,54
Ähnliche Bioeffizienzen werden beobachtet, wenn verschiedene Aquafeeds entweder mit DL-Met oder OH-Met ergänzt werden. Abbildung 4 zeigt, dass ähnliche FCR-Ergebnisse in Studien erzielt wurden, die sich mit der Ergänzung von Fisch- oder Garnelenfutter mit verschiedenen Met-Formen befassten. Trotz der großen Variabilität zwischen den Studien, was auf die unterschiedlichen experimentellen Einstellungen zurückzuführen ist, Futterformulierung, und Garnelen- oder Fischarten, für beide Met-Formen wurden ähnliche FCR-Werte berichtet.
Abschließend, Es gibt wissenschaftliche Beweise dafür, dass OH-Met von aquatischen Arten als Vorläufer von L-Met effizient genutzt werden kann. Es ist wichtig, die strukturellen Unterschiede zwischen DL-Met und OH-Met anzuerkennen, die zu unterschiedlichen Mechanismen für die Absorption und Nutzung dieser beiden Met-Quellen führen. und dass ihre Biowirksamkeit basierend auf der Met-Aufnahme verglichen werden sollte. Daher, DL-Met und OH-Met sind ähnlich wirksam wie Met-Ergänzungen für pflanzliche Aquafeeds.
Abschließend, Es gibt wissenschaftliche Beweise dafür, dass OH-Met von aquatischen Arten als Vorläufer von L-Met effizient genutzt werden kann. Es ist wichtig, die strukturellen Unterschiede zwischen DL-Met und OH-Met anzuerkennen, die zu unterschiedlichen Mechanismen für die Absorption und Nutzung dieser beiden Met-Quellen führen. und dass ihre Biowirksamkeit basierend auf der Met-Aufnahme verglichen werden sollte. Daher, DL-Met und OH-Met sind ähnlich wirksam wie Met-Ergänzungen für pflanzliche Aquafeeds.